


Initialising ...
Initialising ...
Initialising ...
Initialising ...
Initialising ...
Initialising ...
Initialising ...
由良 敬; 郷 通子*
no journal, ,
植物のオルガネラでは、DNAからmRNAへの転写後に、mRNAの塩基が酵素によって変換される場合がある。RNAエディティングとよばれるこの現象では、大部分のRNAエディティングは、mRNA上に一見ランダムに存在し、タンパク質の機能部位とは関係がないアミノ酸残基を編集する。このことからRNAエディティングは、一部の例外を除いて、タンパク質の機能には影響がないと考えられている。そこでわれわれは、今までに報告されているRNAエディティング部位を集め、mRNAとタンパク質のどのような部位が編集されるかを調べた。データを収集した結果、植物のオルガネラでは、365遺伝子中に1923部位のRNAエディティングを見いだすことができた。タンパク質の立体構造がわかっている場合(755部位)において、RNAエディティングによって変化するアミノ酸残基は、タンパク質立体構造形成に重要なヘリックスやコア構造を構成アミノ酸残基である場合が非常に多いことが判明した。タンパク質は立体構造を形成して初めて機能することより、このことはRNAエディティングが立体構造を通してタンパク質の機能に間接的に影響を及ぼしていることを意味する。
佐藤 勝也; 鳴海 一成; 大庭 寛史
no journal, ,
PprMタンパク質はcold shock protein(CSP)のホモログである。大腸菌は9つのCSPを持つが、PprMはラジオデュランスの唯一のCSPであり、大腸菌CspDと進化的に近い。本研究では、放射線応答におけるPprMタンパク質の機能を解析するために、PprMタンパク質を精製し、PprMタンパク質抗体を作成した。この抗体を用いて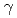 線照射後のPprMタンパク質の細胞内変動を解析したところ、野生株では、線照射,非照射にかかわらず、PprMタンパク質の細胞内発現量は一定であり、
線照射後のPprMタンパク質の細胞内変動を解析したところ、野生株では、線照射,非照射にかかわらず、PprMタンパク質の細胞内発現量は一定であり、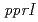 遺伝子破壊株においても発現量は一定であった。しかし、野生株では遺伝子破壊株に比べ、PprMタンパク質のpIが変化していた。さらに、PprMタンパク質は細胞内で多量体を形成していることが明らかになった。PprMタンパク質は大腸菌CspDと比べるとC末端が11アミノ酸残基長く、この領域がラジオデュランスの放射線応答メカニズムに関与していると考えられた。
遺伝子破壊株においても発現量は一定であった。しかし、野生株では遺伝子破壊株に比べ、PprMタンパク質のpIが変化していた。さらに、PprMタンパク質は細胞内で多量体を形成していることが明らかになった。PprMタンパク質は大腸菌CspDと比べるとC末端が11アミノ酸残基長く、この領域がラジオデュランスの放射線応答メカニズムに関与していると考えられた。
玉田 太郎; 新井 栄揮; 正山 祥生; 本庄 栄二郎; 黒木 良太
no journal, ,
タンパク質の立体構造情報が集積される一方で、受容体のような膜タンパク質の構造情報は遅れがちである。細胞の表層に発現する受容体は、創薬における重要な標的であることが知られるが、その多くは一般的に不安定でプロテアーゼ消化されやすく、付加した糖鎖による不均一性から構造解析の対象となりにくかった。そこでわれわれはこれらのタンパク質を効果的に結晶化させるため、抗体、特にFabフラグメントとの共結晶化研究を行っている。Fabとの共結晶化では、複合体化による目的タンパク質の安定化,Fab分子による目的タンパク質の結晶化促進,既知のFab構造の位相情報を利用した迅速な構造決定などが期待できる。さらに抗体自身は医薬品や診断薬としての重要な用途があるため、抗原のエピトープ決定や抗体の高機能化などにおいて産業応用面においても重要な研究である。本発表ではわれわれの抗体を用いた結晶化研究で得られたさまざまな知見について紹介する。
安達 基泰; 玉田 太郎; 佐藤 勝也; 由良 敬; 鳴海 一成; 黒木 良太
no journal, ,
PprAはデイノコッカスより単離された新規なDNA修復促進蛋白質であり、高度放射線抵抗性において最も重要な役割を担っている。本研究では、PprAの構造と機能の関係を解明することを目的として、PprAとDNAとの相互作用解析を行った。大腸菌発現系を用いて調製した組換えPprAを精製し、DNAとの相互作用をアガロースゲルシフトアッセイとゲル濾過により検討した。その結果、PprAは1分子の2本鎖DNA(pUC19,2686 bp)に少なくとも約280分子の結合が可能であること、PprAとDNAとの複合体形成が、1mMの低濃度のMg, CaあるいはSrイオンにより促進されること、PprAの多量体構造の形成及びPprAとDNAとの複合体の形成の両方に塩濃度依存性(0-0.4M、酢酸ナトリウム)があることが示された。また、直鎖状の2本鎖DNAとPprAが複合体を形成する場合、PprAの濃度上昇に伴って複合体どうしが会合し、大きな分子量を持つ会合体を形成することを明らかにした。この結果は、放射線によって切断されたゲノムDNAの2つの末端どうしをPprAが近づける役割を持つことを示唆し、PprAの機能発現において重要な知見であると考えられる。
佐藤 勝也; 大庭 寛史; Sghaier, H.*; 鳴海 一成
no journal, ,
放射線抵抗性細菌デイノコッカス・ラジオデュランスのLexA2蛋白質の放射線応答機構における役割を明らかにするために、遺伝子破壊株を作製し、線に対する感受性を調べるとともに、照射前後での放射線誘導性DNA修復遺伝子の発現誘導と蛋白質の細胞内量変動を解析した。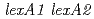 遺伝子二重破壊株では、野生株と同様に線照射後にRecA蛋白質の細胞内量が増加したことから、LexA1及びLexA2蛋白質はどちらもRecA蛋白質の誘導には関与していないことが明らかになった。興味深いことに、
遺伝子二重破壊株では、野生株と同様に線照射後にRecA蛋白質の細胞内量が増加したことから、LexA1及びLexA2蛋白質はどちらもRecA蛋白質の誘導には関与していないことが明らかになった。興味深いことに、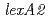 遺伝子破壊株及び遺伝子二重破壊株は、野生株よりも線に対して耐性を示した。遺伝子破壊株では、ラジオデュランスに特有の放射線誘導性DNA修復促進蛋白質PprAの遺伝子発現に必要なプロモーター活性の促進が観察されたことから、LexA2蛋白質には通常の細胞増殖等に不利益となるPprA蛋白質の過剰な細胞内蓄積を抑制する機能があると考えられた。以上のことから、遺伝子破壊株で示された高い線耐性の一因は、放射線照射後にPprA蛋白質の機能が野生株以上に増強されることによると考えられた。
遺伝子破壊株及び遺伝子二重破壊株は、野生株よりも線に対して耐性を示した。遺伝子破壊株では、ラジオデュランスに特有の放射線誘導性DNA修復促進蛋白質PprAの遺伝子発現に必要なプロモーター活性の促進が観察されたことから、LexA2蛋白質には通常の細胞増殖等に不利益となるPprA蛋白質の過剰な細胞内蓄積を抑制する機能があると考えられた。以上のことから、遺伝子破壊株で示された高い線耐性の一因は、放射線照射後にPprA蛋白質の機能が野生株以上に増強されることによると考えられた。
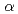 1鎖及びインターロイキン-4受容体鎖細胞外ドメインの調製と性質
1鎖及びインターロイキン-4受容体鎖細胞外ドメインの調製と性質本庄 栄二郎; 正山 祥生; 玉田 太郎; 有馬 和彦*; 金地 佐千子*; 出原 賢治*; 黒木 良太
no journal, ,
アレルギーの発症に関与するサイトカインIL-13の受容体であるIL-13R1及びIL-4とリガンドであるIL-13との相互作用を解析するために、各受容体の細胞外ドメインの発現と調製を行った。IL-13R1及びIL-4の細胞外ドメインを、目的蛋白質の細胞外領域をアミノ酸配列の相同性と立体構造の予測モデルによって絞り込み、その領域を、トロンビン切断部位を有するポリアラニンリンカーを介し抗体Fc領域と融合した形態で発現させた。タンパク質の発現には片倉工業のトランスファーベクター/カイコ発現の系を利用した。発現されたFc融合蛋白質は、プロテインAカラムによって迅速に精製でき、IL-13R1-Fcは、カイコ体液1mlあたり0.5mg、IL-4R-Fcは、0.1mg調製した。最終的に、融合したFc領域をプロテアーゼ消化によって除去し、目的受容体の細胞外領域を調製した。調製した受容体のリガンド結合能を評価したところ、IL-4Rの存在下において、IL-13リガンドのIL-13R1への親和性が増大することが確認できた。
中川 繭; 坂本 綾子; 高橋 真哉*; 田中 淳; 鳴海 一成
no journal, ,
われわれはシロイヌナズナのUV感受性変異体の原因遺伝子として、損傷乗り越え複製(TLS)型DNAポリメラーゼ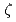 のサブユニットであるREV3の相同遺伝子AtREV3を単離した(Sakamoto et al. 2003)。さらにdCMPトランスフェラーゼ活性を示すREV1のシロイヌナズナの相同遺伝子AtREV1を単離した(Takahashi et al. 2005)。これらのREV相同遺伝子の欠損は植物体の生存には影響しないが、DNA複製に関与することが推測され、UVのほか、線やDNA架橋剤MMCなどの変異原に対して感受性を示した。本研究では、ナンセンスコドンとなる点変異を導入したuidA遺伝子をリポーターとして用いて、塩基置換型復帰変異を指標にUV誘発変異頻度の解析を行った。rev3, rev1欠損植物体のUV照射及び線照射による突然変異の頻度は、ともに野生型に比べ著しい減少を示した。この結果から、AtREV3及びAtREV1の欠損植物体では、正常なTLSが機能しないため、UV, 線感受性になっているという可能性が示唆された。
のサブユニットであるREV3の相同遺伝子AtREV3を単離した(Sakamoto et al. 2003)。さらにdCMPトランスフェラーゼ活性を示すREV1のシロイヌナズナの相同遺伝子AtREV1を単離した(Takahashi et al. 2005)。これらのREV相同遺伝子の欠損は植物体の生存には影響しないが、DNA複製に関与することが推測され、UVのほか、線やDNA架橋剤MMCなどの変異原に対して感受性を示した。本研究では、ナンセンスコドンとなる点変異を導入したuidA遺伝子をリポーターとして用いて、塩基置換型復帰変異を指標にUV誘発変異頻度の解析を行った。rev3, rev1欠損植物体のUV照射及び線照射による突然変異の頻度は、ともに野生型に比べ著しい減少を示した。この結果から、AtREV3及びAtREV1の欠損植物体では、正常なTLSが機能しないため、UV, 線感受性になっているという可能性が示唆された。
とREV1蛋白質の解析坂本 綾子; 高橋 真哉*; 岩井 成憲*; 清水 喜久雄*
no journal, ,
われわれはシロイヌナズナのDNAポリメラーゼ及びREV1蛋白質の遺伝子を単離し、解析を行ってきた。DNAポリメラーゼのサブユニットをコードする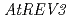 ,
, 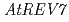 、及び
、及び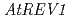 遺伝子を欠損したシロイヌナズナは、通常の生育条件下では正常に生長するが、紫外線照射下で生育させると、野生型に比べて強い生長阻害がみられた。また、播種後3日目の幼植物体に対して紫外線や線、及びクロスリンク試薬などを与えると、及び欠損株では、野生型に比べて根端の分裂組織の増殖が阻害され、根の伸長が抑制されるという結果になった。このことから、シロイヌナズナのDNAポリメラーゼ及びREV1蛋白質が、DNA損傷による細胞増殖の停止を忌避する働きをもっていることが予想された。そこで、AtREV1蛋白質の機能を明らかにする目的で
遺伝子を欠損したシロイヌナズナは、通常の生育条件下では正常に生長するが、紫外線照射下で生育させると、野生型に比べて強い生長阻害がみられた。また、播種後3日目の幼植物体に対して紫外線や線、及びクロスリンク試薬などを与えると、及び欠損株では、野生型に比べて根端の分裂組織の増殖が阻害され、根の伸長が抑制されるという結果になった。このことから、シロイヌナズナのDNAポリメラーゼ及びREV1蛋白質が、DNA損傷による細胞増殖の停止を忌避する働きをもっていることが予想された。そこで、AtREV1蛋白質の機能を明らかにする目的で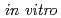 におけるポリメラーゼ活性を解析したところ、AtREV1は脱塩基部位を持つDNAを鋳型として逆鎖のプライマー末端にdCMPを挿入した。このことから、AtREV1蛋白質が脱塩基部位を持つDNAの複製に関与していることが示唆された。一方で、AtREV1蛋白質は、紫外線損傷DNAの逆鎖に対しては塩基挿入活性を示さなかった。このことから、AtREV1は、dCMPトランスフェラーゼ活性のほかに未同定の機能があり、これが失われることにより植物に紫外線感受性が生じたことが示唆された。
におけるポリメラーゼ活性を解析したところ、AtREV1は脱塩基部位を持つDNAを鋳型として逆鎖のプライマー末端にdCMPを挿入した。このことから、AtREV1蛋白質が脱塩基部位を持つDNAの複製に関与していることが示唆された。一方で、AtREV1蛋白質は、紫外線損傷DNAの逆鎖に対しては塩基挿入活性を示さなかった。このことから、AtREV1は、dCMPトランスフェラーゼ活性のほかに未同定の機能があり、これが失われることにより植物に紫外線感受性が生じたことが示唆された。